
The Mysteries of Scorpion Fluorescence
Properties and Fluorescent Compounds
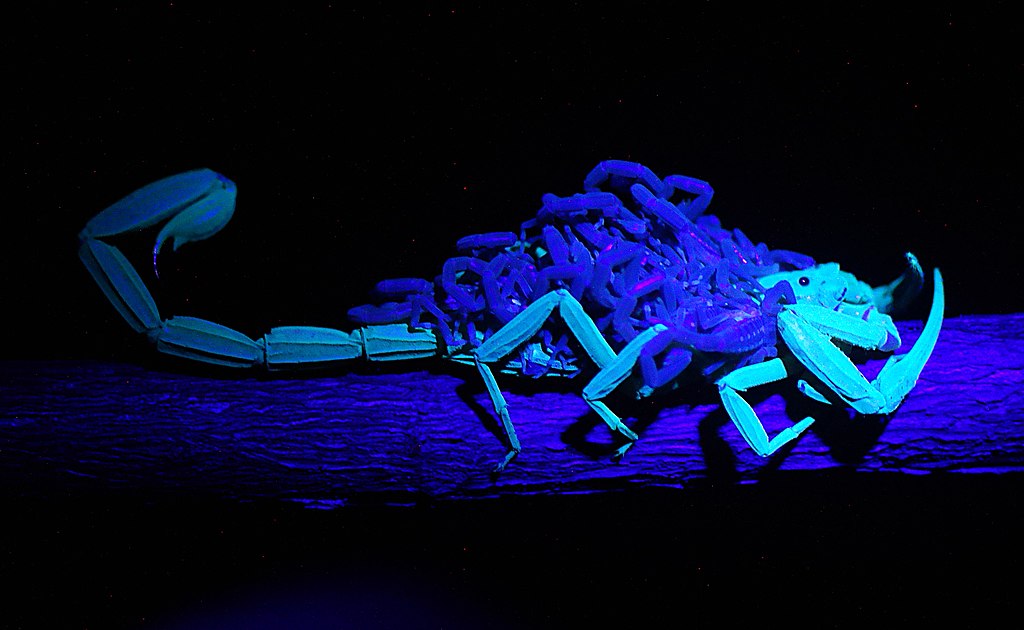
Scorpion fluorescence was first reported in 1954 and has fascinated researchers and enthusiasts ever since (Lawrence, 1954; Frost et al., 2001). Most extant scorpion species are thought to fluoresce under ultraviolet (UV) illumination (see “Non-fluorescent Scorpions” for exceptions).
Characteristic features of the phenomenon include the absence of fluorescence in first-instar juveniles and the weak or absent fluorescence immediately after ecdysis; in contrast, the shed exuviae often exhibit strong fluorescence. Scorpion remains may also fluoresce; under favorable preservation conditions some specimens retain fluorescence for centuries (Fet et al., 2011).
The scorpion fluorescence phenomenon is a form of photoluminescence (Fasel et al., 1997): certain cuticular compounds absorb light at specific wavelengths (excitation) and re-emit light at longer wavelengths. In scorpions, excitation typically occurs near the ultraviolet range (around 350 nm), and emission is usually in the blue–green range (approximately 500 nm).
In the Emperor Scorpion (Pandinus imperator), excitation peaks near 365 nm, while emission peaks around 500 nm (Fasel et al., 1997).


Fluorescent compounds are concentrated in the outermost layer of the cuticle—the hyaline layer—approximately four micrometres thick. Two compounds have been chemically identified from scorpion cuticle: β-carboline (norharman) and a coumarin derivative commonly referred to as hymecromone (4-methyl-7-hydroxycoumarin, 4-methylumbelliferone) (Stachel et al., 1999; Frost et al., 2001).
β-Carboline is an alkaloid found in various plants and animals, while hymecromone (a coumarin derivative) is widespread in plants and some microbes but is rarely reported from animals, and its presence in arthropods is unusual (Frost et al., 2001).


Extracts of β-carboline are excited near 350 nm and emit around 450 nm (Stachel et al., 1999), whereas extracts containing hymecromone have been reported to be excited 409.5 nm and to emit 441 nm (Frost et al., 2001). Differences between these values and observed whole-cuticle spectra may reflect matrix effects in the intact cuticle or combined contributions from multiple compounds.
Functional Hypotheses
What function, if any, does cuticular fluorescence serve for scorpions? Multiple hypotheses have been proposed, drawing on observations of fluorescence in other animal groups and on scorpion sensory biology:
- Detection of ultraviolet (Blass & Gaffin, 2008)
- Detection of moonlight (Kloock et al., 2010)
- Mate or species recognition
- Deterrence of predators or competitors
- Mate attraction
- Prey attraction or facilitation of prey capture

Scorpion photoreceptors show sensitivity near 500 nm (Machan, 1968), which overlaps with the emission range of cuticular fluorescence; this overlap leaves open the possibility of visual communication mediated by fluorescence. Additionally, scorpion cuticle itself may mediate UV perception via cuticular photoreception (Blass & Gaffin, 2008; Kloock et al., 2010), potentially linking fluorescence and UV responsiveness.
Conversely, some authors argue that fluorescence may be functionally neutral. In plants, coumarins are known to function as UV screening compounds. Similarly, it is possible that hymecromone in scorpions once served a comparable photoprotective role. Although most extant scorpions are now nocturnal, they may still synthesize hymecromone as an evolutionary remnant of this ancestral UV-shielding mechanism (Frost et al., 2001). The absence of fluorescence in first-instar juveniles and immediately post-ecdysis individuals suggests that fluorescence could sometimes be a by-product of cuticle maturation rather than an adaptive signal (Stachel et al., 1999).
At present there is no universally accepted explanation; the functional significance of scorpion fluorescence remains an open research question.
Evolutionary Context
Chemical analyses and cuticular cross-section studies show that fluorescent compounds in scorpions are located in the hyaline layer of the cuticle. Recent comparative work suggests that cuticular photoluminescence may be widespread among chelicerates.

Rubin et al. (2017) reported that UV-induced fluorescence is detectable across multiple arachnid groups, but fluorescence outside Scorpiones often localizes to exposed soft tissues (e.g., membranous joints) and likely involves different compounds than those in scorpion cuticle. Horseshoe crabs (marine chelicerates) also exhibit cuticular fluorescence and possess a hyaline layer analogous to that of scorpions, though their fluorescent compounds remain to be fully characterized (Rubin et al., 2017). Fossil eurypterid cuticle shows hyaline-like layers as well, suggesting that cuticular fluorescence might be plesiomorphic within Chelicerata.
If cuticular fluorescence originated in aquatic chelicerate ancestors, its persistence in terrestrial scorpions could be nonadaptive in current contexts—i.e., a retained ancestral trait with limited contemporary function.

Non-fluorescent Scorpions
Although UV fluorescence has been observed in many scorpion taxa and was long considered universal, not all described species have been tested. In 2012, Lourenço reported several species in the family Chaerilidae that do not fluoresce under UV illumination (Lourenço, 2012).
Chaerilids are a modestly sized group (about fifty described species) distributed from India to Southeast Asia. Studies of nine chaerilid species found no fluorescence regardless of eye presence or habitat (surface vs. cave), suggesting a lineage-wide loss or absence of cuticular fluorescence rather than a simple ecological loss of function.
Photographs of non-fluorescent chaerilids are available in the cited literature and online resource.
The occurrence of non-fluorescent scorpions demonstrates that fluorescence is not an absolute diagnostic trait of Scorpiones and supports the view that fluorescence may be evolutionarily labile or sometimes functionally redundant.
References ▼
- Frost, L.M., Butler, D.R., O'Dell, B., & Fet, V. (2001). A coumarin as a fluorescent compound in scorpion cuticle. In V. Fet & P.A. Selden (Eds.), Scorpions 2001: In Memoriam Gary A. Polis (pp. 365–368). British Arachnological Society.
- Lawrence, R.F. (1954). Fluorescence in Arthropoda. Journal of the Entomological Society of Southern Africa, 17, 167–170.
- Fet, V., Soleglad, M.E., & Zonstein, S.L. (2011). The genus Akrav Levy, 2007 (Scorpiones: Akravidae) revisited. Euscorpius, 134, 1–49.
- Fasel, A., Müller, P.-A., Suppan, P., & Vauthey, E. (1997). Photoluminescence of the African scorpion Pandinus imperator. Journal of Photochemistry and Photobiology B: Biology, 39, 96–98.
- Stachel, S.J., Stockwell, S.A., & Van Vranken, D.L. (1999). The fluorescence of scorpions and cataractogenesis. Chemistry & Biology, 6, 531–539.
- Blass, G.R.C., & Gaffin, D.D. (2008). Light wavelength biases of scorpions. Animal Behaviour, 76, 365–373.
- Kloock, C.T., Kubli, A., & Reynolds, R. (2010). Ultraviolet light detection: A function of scorpion fluorescence. Journal of Arachnology, 38, 441–445.
- Machan, L. (1968). Spectral sensitivity of scorpion eyes and the possible role of shielding pigment effect. Journal of Experimental Biology, 49, 95–105.
- Rubin, M., Lamsdell, J.C., Prendini, L., & Hopkins, M.J. (2017). Exocuticular hyaline layer of sea scorpions and horseshoe crabs suggests cuticular fluorescence is plesiomorphic in chelicerates. Journal of Zoology, 303(4), 245–253.
- Lourenço, W.R. (2012). Fluorescence in scorpions under UV light; Can chaerilids be a possible exception? Comptes Rendus Biologies, 335, 731–734.
