
サソリの光るなぞ (蛍光現象)
蛍光の特徴と蛍光物質
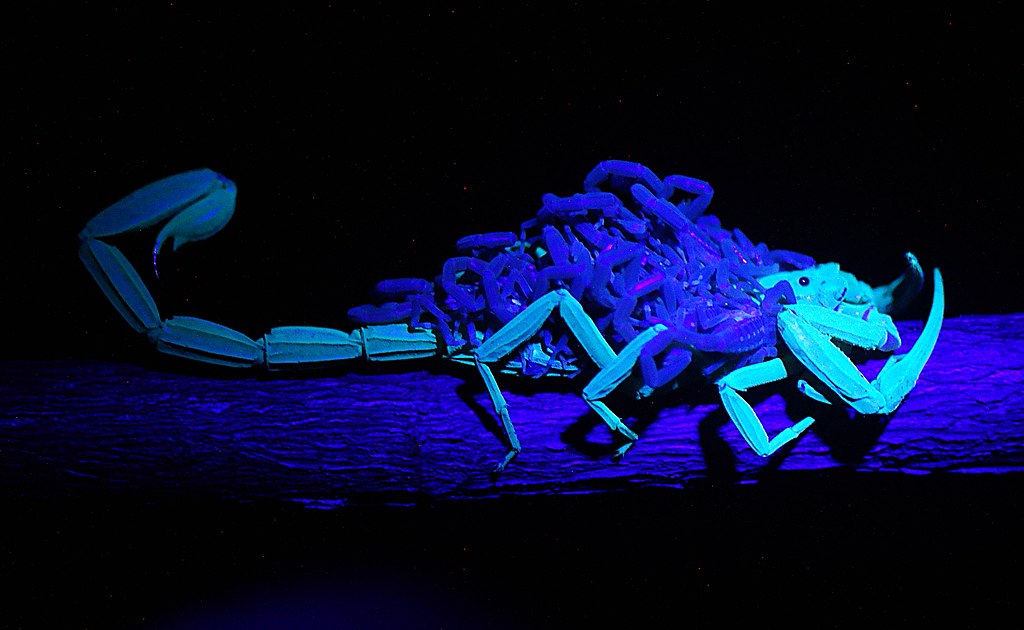
サソリの蛍光現象は1954年に初めて報告されて以来 (Frost et al., 2001; Lawrence, 1954)、多くのサソリ研究者や愛好家を魅了してきました。現生サソリ目のほぼ全ての種が蛍光すると考えられています (例外については「蛍光しないサソリ」をご覧ください)。
蛍光反応の特徴として、2齢以降のサソリは蛍光しますが、1齢若虫では蛍光しません。また、脱皮直後のサソリも蛍光しないか、非常に蛍光が弱いのに対し、残された脱皮殻は蛍光反応を示します。
さらに、サソリの死骸でも蛍光し、保存状態が良ければ、200年経っても蛍光反応を示すそうです (Fet et al., 2011)。
サソリの蛍光現象はフォトルミネセンス (photoluminescence; Fasel et al., 1997) と呼ばれ、物質がある波長の光を吸収した後、別の波長の光を放出 (発光) する現象です。単純な反射とは異なります。光を吸収する過程を「励起 (れいき)」と言い、吸収する波長の光を「励起光」と呼びます。
サソリの蛍光反応の場合、約350nmの紫外線 (紫外光) によって励起され、約500nmの青色~緑色の光を放出します。
ダイオウサソリ (Pandinus imperator) では励起光の最大波長が365nm、放出される (蛍光する) 波長の最大値が約500nmとなるようです (Fasel et al., 1997)。


サソリの蛍光物質は、外骨格 (外皮) の最外層であるヒアリン層 (約4μmの厚さの部分) に集中しているそうです。
現在までのところ、蛍光物質としてはβ-カルボリン (β-carboline) とヒメクロモン (Hymecromone) という2つが確認 (同定) されています (Frost et al., 2001; Stachel et al., 1999)。
β-カルボリン (別名:norharman) はアルカロイド (窒素原子を含み、アルカリ性を示す有機化合物で、多くは有毒) であり、様々な植物や動物でみられる物質だそうです。
一方、ヒメクロモン (別名:4-methyl-7-hydroxycoumarin、4-methylumbelliferone) はクマリン (coumarin) という化合物の一種であり、植物に代表される成分で、菌類やバクテリアにも含まれますが、動物では非常に珍しく、節足動物ではこれまで発見されていないそうです (Frost et al., 2001)。


β-カルボリンのアルコール抽出液は約350nmの波長で励起され、約450nmの蛍光を放出するそうです (Stachel et al., 1999)。また、ヒメクロモンの抽出液は409.5nmの波長で励起され、441nmの蛍光を放出するそうです (Frost et al., 2001)。
サソリの蛍光と波長が異なりますが、これはサソリの外皮に存在する状態と抽出液とでの違いによるものであったり、2種類の化合物の反応が合わさった結果かもしれません。
蛍光の機能的役割
蛍光現象は、サソリにとってどのような意味をもっているのでしょうか。
蛍光現象はサソリ以外の動物でもみられ、交配・繁殖のためなど、機能的な意味をもっている場合もあるようです。
サソリの蛍光現象の機能的役割については、これまでにいくつもの説が提唱されています。
- 紫外線の感知 (Blass and Gaffin, 2008)
- 月光の感知 (Kloock et al., 2010)
- 異性や種を区別するため
- 天敵やライバルを寄せつけないため
- 異性を惹きつけるため
- 獲物を引き寄せる・捕獲のため

これまでに、サソリの視覚は約500nmの緑色の光に対して感受性があることがわかっており (Machan, 1968)、サソリどうしで蛍光によってコミュニケーションをとっている可能性は十分にあります。
また、サソリの外骨格 (外皮) は紫外線を感知できるとも考えられているため (Blass and Gaffin, 2008; Kloock et al., 2010)、蛍光反応は紫外線に対する反応に関係している可能性もあります。
反対に、サソリの蛍光には特に機能的な意味が無いという説もあります。
植物ではクマリンが紫外線を防ぐ役割をしている場合があるそうで、サソリにおいてもかつてはヒメクロモンが日よけの役割を担っていたのが、現在では夜行性の種が多くなったにもかかわらず、日よけのメカニズムの名残としてヒメクロモンを合成しているだけかもしれません (Frost et al., 2001)。
また、1齢若虫や脱皮直後のサソリでは蛍光反応を示さないため、蛍光物質は、外骨格が硬化するための反応のなかで副産物として合成されているだけなのかもしれません (Stachel et al., 1999)。
いずれにしても、蛍光反応についてはいまだに決定的な説が無く、蛍光現象の意味が明らかにされるのはまだまだ先かもしれません。
蛍光現象の進化的位置づけ
サソリの蛍光については、β-カルボリンとヒメクロモンが蛍光物質として同定されており、これらの蛍光物質は外骨格 (クチクラ) のヒアリン層 (hyaline layer) という部分に存在することが知られています。

Rubin et al. (2017) によると、紫外光による蛍光自体は全ての蛛形類で確認されたのだそうですが、サソリ類以外の蛛形類で見られる蛍光は、外骨格で覆われていない外皮部分 (外骨格の間の関節部分など) で見られ、蛍光物質もサソリの外骨格のものとは異なるようです。
一方、水生鋏角類であるカブトガニ類では、サソリ類と同様、外骨格の蛍光が確認されました (Rubin et al., 2017)。
さらに、外骨格の断面を観察したところ、サソリ類と同じように、ヒアリン層が確認されたそうです (蛍光物質の同定はまだされていません)。また、ウミサソリの化石でも、蛍光はしませんが、ヒアリン層が確認できたそうです (Rubin et al., 2017)。
ここから、サソリ類で見られる蛍光現象は、水生鋏角類の祖先に由来する可能性が出てきます。
そして、もし蛍光現象が水生の祖先に由来するのであれば、陸上生活のサソリにおいては蛍光現象は特に意味をもたないのかもしれません。

蛍光しないサソリ
紫外線による蛍光はこれまで多くのサソリで確認され、全てのサソリで共通する現象だと考えられてきました。
しかし、当然ですが、これまでに記載された全ての種で蛍光が調べられたわけではありません。
2012年になって、紫外線に対して蛍光反応を示さないサソリが報告されました (Lourenço, 2012)。
アゴハリサソリ科 (Chaerilidae) は、インドから東南アジアにかけて生息しており、約50種からなる小さなグループですが、このアゴハリサソリ科のサソリでは紫外線に対して蛍光反応を示さないそうです。
蛍光しないサソリの写真 (外部サイト)
アゴハリサソリ科は、洞窟性のサソリや雨林にすむサソリ、山岳地域の森林に生息するサソリなど、種によって生息環境が様々ですが、これらのサソリ9種で蛍光反応を調べた結果、眼の有無や生息環境の違いに関係なく、9種全てで蛍光が見られなかったそうです (Lourenço, 2012)。
眼を失ったサソリや洞窟性のサソリなど、サソリどうしで蛍光反応を見分けられない状況であれば、蛍光する必要がないため、蛍光しなことが環境への適応と考えられます。しかし、アゴハリサソリ科のサソリが蛍光しないことは紫外線が入らない環境への適応などではなく、アゴハリサソリ科が系統的に蛍光反応を失った (もしくは蛍光しないことを獲得した) と考えられます。
アゴハリサソリ科で蛍光反応を示さない理由はわかっていませんが、少なくともアゴハリサソリ科のサソリは蛍光反応を示さなくても生きていけるわけで、もしかすると、上述のようにサソリにおいては蛍光現象は特に機能的な意味をもっていないのかもしれません。
出典 ▼
- Frost, LM., DR. Butler, B. O'Dell and V. Fet (2001) A coumarin as a fluorescent compound in scorpion cuticle. In Fet, V. and P.A. Selden (eds.), Scorpions 2001: In Memoriam, Gary A. Polis, British Arachnological Society, Buckinghamshire, pp.365-368.
- Lawrence, RF. (1954) Fluorescence in Arthropoda, Journal of the Entomological Society of South Africa, 17:167-170.
- Fet, V., ME. Soleglad and SL. Zonstein (2011) The genus Akrav Levy, 2007 (Scorpiones: Akravidae) revisited, Euscorpius, 134:1-49.
- Fasel, A., P-A. Muller, P. Suppan and E. Vauthey (1997) Photoluminescence of the African scorpion "Pandinus imperator", Journal of Photochemistry and Photobiology. B, Biology, 39:96-98.
- Stachel, SJ., SA. Stockwell, and DL. Van Vranken (1999) The fluorescence of scorpions and cataractogenesis, Chemistry and Biology, 6:531-539.
- Blass, GRC. and DD. Gaffin (2008) Light wavelength biases of scorpions, Animal Behaviour, 76:365-373.
- Kloock, CT., A. Kubli and R. Reynolds (2010) Ultraviolet light detection: a function of scorpion fluorescence, Journal of Arachnology, 38:441-445.
- Machan, L. (1968) Spectral sensitivity of scorpion eyes and the possible role of shielding pigment effect, The Journal of Experimental Biology, 49:95-105.
- M. Rubin, JC. Lamsdell, L. Prendini and MJ. Hopkins (2017) Exocuticular hyaline layer of sea scorpions and horseshoe crabs suggests cuticular fluorescence is plesiomorphic in chelicerates, Journal of Zoology, 303(4):245-253.
- Lourenço, WR. (2012) Fluorescence in scorpions under UV light; can chaerilids be a possible exception?, Comptes Rendus Biologies, 335:731-734.
