
マダラサソリは外来種なのか?
4.マダラサソリは外来種?在来種?
ここからは、第3章でまとめた外来種か在来種かを判定する5つのポイントについて、それぞれマダラサソリで見ていきます。
i.マダラサソリについての記録
Isometrus maculatusという種に対して「マダラサソリ」という和名が命名されたのは1906年であり (岩川, 1906)、(それまでは種名がついていなかったため) 日本国内におけるマダラサソリの記録もこの文献が最古ではないかと思われます。岩川 (1906) は「本種は、前種 (筆者注:ヤエヤマサソリ) の如く多數ならざるが如きも、亦八重山群島に産し、今回余の目撃せる本種の標本は、博物館と大學に藏せる唯二尾のみなり。」とし、八重山諸島で採集された標本を確認しています。標本の採集時期は1900年前後からそれ以前と思われますが、記述はありません。
日本国内のサソリについての岩川 (1906) より古い記録として、黒岩 (1899a) による記述が確認できました。黒岩は1898年~1899年に石垣島を訪れています (黒岩, 1899b)。しかし、「蝎及サソリモドキの如きは、殆本島固有なり」とあり、サソリモドキと併記されていることから、サソリモドキと同じような場所でサソリを発見した可能性があり、これはヤエヤマサソリを指している可能性もあります。
マダラサソリという和名は付いていなかったものの、先島諸島内でいつから知られていたかを調べるには、現地の郷土資料を調べる必要があると思いますが、これはなかなか大変なことだと思います。
江戸時代には国外から来た商船中でサソリが見つかった例が知られていますが、当時の文献 (スケッチ) のうち、高島 (1943) によってマダラサソリと述べられているものもあります。このような事情から、マダラサソリが江戸時代に日本国内へ導入された可能性もありますが、こちらも不明です。
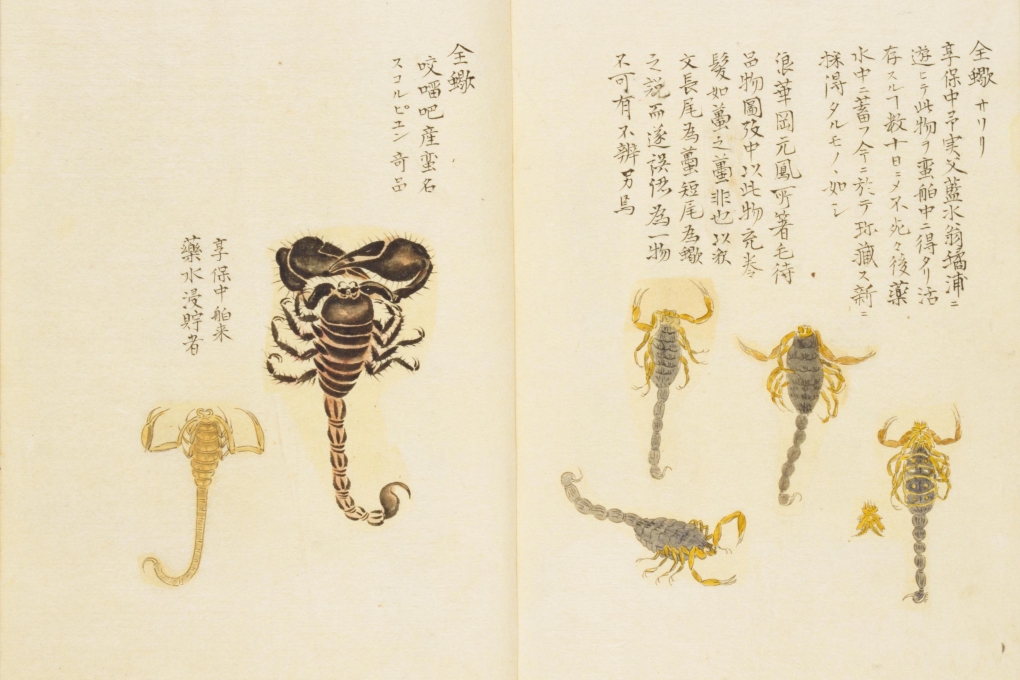
高島 (1943) は左の個体をマダラサソリのオスとしている。
国内で唯一、小笠原諸島に分布する個体群は外来個体群だと考えられています。
高島 (1954) や古川・高島 (1942) は、1931年頃から父島・母島でマダラサソリが見られるようになったとしており、サイパン方面から輸入したバナナに紛れてきたと考えられています。その後、父島・母島では、マダラサソリとオオムカデの駆除目的でオオヒキガエルが導入され、マダラサソリは見られなくなりました (現在ではオオヒキガエルが外来種として問題になっています:環境省, 2007)。
導入時期や経路は父島・母島とは異なると思われますが、現在は、小笠原諸島では硫黄島のみでマダラサソリが分布しているとされています (新たな外来種の侵入・拡散防止に関するワーキンググループ, 2013; 久保, 2001)。
マダラサソリの記録:
現在のところ1906年が最古。起源 (導入の有無等) は不明。
ii.マダラサソリの考古学的記録
現在のところ、Isometerus属と考えられる化石の記録は見つかりませんでした。
iii.マダラサソリの遺伝的解析
国立遺伝学研究所のDDBJにて塩基配列データを調べてみましたが、マダラサソリの分布について遺伝的解析を行った、生物系統地理学や分子系統解析といった研究は今のところ見つかりませんでした。
iv.マダラサソリの分布
日本国内 (外来個体群とされる小笠原諸島は省略) でマダラサソリの生息・発見の記録を、文献およびインターネット情報で調べたところ、以下のようになりました (インターネットの情報は個人のブログ・SNSの場合もあり、不正確な可能性もあるため、出典は控えます)。
- 宮古島 (相原・秋山, 2007; 青木, 2015; 下謝名, 1972)
- 伊良部島 (下謝名, 1972)
- 下地島
- 池間島
- 多良間島
- 石垣島 (相原・秋山, 2007; 青木, 2015; 下謝名, 1972)
- 西表島 (相原・秋山, 2007; 青木, 2015)
- 新城島 (上地島:山﨑ら, 2016)
- 黒島
- 波照間島
- 与那国島 (相原・秋山, 2007)
また、日本周辺・アジアでマダラサソリと、近縁なIsometrus属のサソリの分布を調べた結果は以下のようになります (各国内の細かな地域は省略)。
- マダラサソリ:インド・パキスタン・モルディブ・スリランカ・カンボジア・インドネシア・ラオス・マレーシア・ミャンマー・フィリピン・タイ・ベトナム・パプアニューギニア (Fet et al., 2000)、台湾 (Di et al., 2014)、中国 (海南島:Di et al., 2013)
- I. amboli:インド (Sulakhe et al., 2020)
- I. formosus:インドネシア (ジャワ島)・シンガポール (Kovařík, 2003)
- I. tamhini:インド (Sulakhe et al., 2020)
- I. thurstoni:インド (Kovařík, 2003; Kovařík et al., 2016)
- I. thwaitesi:スリランカ (Kovařík, 2003; Kovařík et al., 2016)

黄緑:マダラサソリ、オレンジ:マダラサソリと近縁種。
図のように、マダラサソリは飛び石的な分布ではなく、先島諸島全域に分布しているようです。互いに数十キロメートル離れた各島に別々に導入されたと考えるのは難しいかもしれません。しかし、この地域も古くから人が住んでいたことが知られており (最古の記録は約28,000年前:琉球大学, 2019)、江戸時代以前から海外との交流もあったと思われ、人や物資の移動に伴って、偶然 (非意図的) にマダラサソリが導入された可能性もあります。
Lourenço and Huber (2002) は、スリランカではマダラサソリが唯一内陸部で野生下で見つかるため、元々の生息地 (original area of distribution) はスリランカかもしれないと述べています。これに対し、Kovařík et al. (2016) は、この主張は根拠の無い、単なる憶測だと反論しています。ただし、マダラサソリに近縁なI. thurstoniとI. thwaitesiが生息しているため、インドとスリランカはマダラサソリが元々分布していた (indigenous) 可能性があるとしています (Kovařík et al., 2016)。
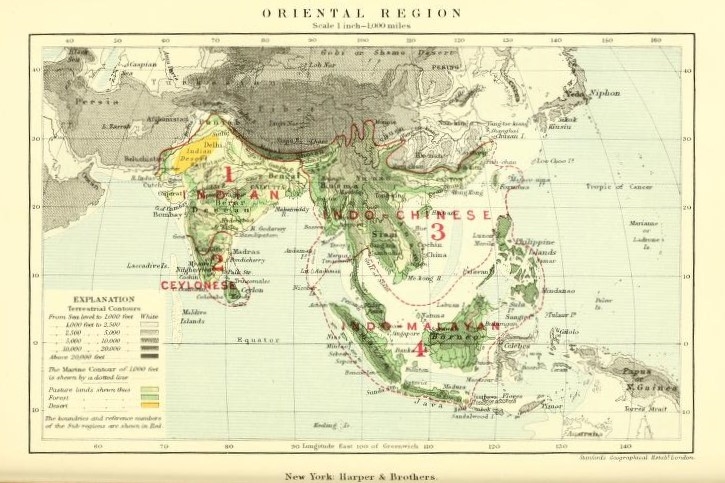
Pocock (1900) は、マダラサソリについて、東洋区から船舶によってアジア・アフリカ・アメリカの熱帯・亜熱帯へ導入されたと述べています。ここでの東洋区 (Oriental Region) とは、Wallace (1876) の提唱した動物地理区のひとつであり、先島諸島もその範囲内にあると考えられます。ただし、マダラサソリの東洋区起源説について、根拠は述べられていません (東洋区を起源とすると明言もしていません)。
また、マダラサソリについて報告されている多くの地域は人為的に分布したと考えられていますが (Pocock, 1900; Fet et al., 2000; Kovařík et al., 2016)、Kovařík et al. (2016) によると、発見の報告が最も多かった1940年頃までは木造の船と木造の建物が多かったことが原因であり、建築材料や移動手段の変化によって、多くの場所では報告が全くなくなったそうです。よって、導入が一時的な場合、過去に報告があっても現在まで個体群を維持している地域は少ないと考えています (Kovařík et al., 2016)。この考えを基にすると、少なくとも1900年頃から現在までの120年間個体群を維持している日本のマダラサソリは、導入よりも分散によって先島諸島へ分布を広げた可能性が十分にあるかもしれませんが、どうでしょうか。
以上を総合的に考えると、少なくともマダラサソリは南アジアや東南アジアのほうから先島諸島へ入ってきたと考えられますが、もし、先島諸島の分布が、導入でなく、分散によって分布を拡大したのであれば、どのような方法が考えられるでしょうか。
a.陸上を移動
未だ議論のあるところではありますが、鹿児島の薩南諸島を含む南西諸島 (琉球弧) は、氷期に海面が低下することによって、過去何度か台湾や大陸 (中国) などと陸続き (陸橋) であった時期があり、その時期に大陸や南方から動植物が渡来したと考えられています (木村, 1996; 諸喜田, 1996; 安間, 2001; 河村, 2014)。
また、琉球弧上には、2か所で水深1,000mを超す凹みができており、それぞれトカラギャップ (トカラ海裂)・ケラマギャップ (ケラマ海裂) と呼ばれています (木村, 1996)。
海面が下がって陸地が拡大した時期でも、トカラギャップ・ケラマギャップがあるために、陸地が完全にはつながらない時期もあったと考えられており、トカラギャップ・ケラマギャップを境に地理的に分断された3つの地域で、生物の分布や特徴も「北琉球」「中琉球」「南琉球」と3つのグループに分かれることがいくつかの生物で知られています (諸喜田, 1996; 宮城ら, 2002; 環境省, 2010)。

上述のように、日本国内のマダラサソリは先島諸島全域に分布している可能性があり、南琉球の区域に一致します。人為的に導入されたのであれば、宮古諸島より北 (中琉球) で分布していてもおかしくないと思うのですが、もし陸橋を渡って日本国内に分布を広げたのであれば、ケラマギャップによって南琉球と中琉球が隔てられていた時期に渡来したと考えられなくもないでしょう (ホントに単なる偶然かもしれませんし、南琉球と中琉球で何かしら環境の違いがあったのかもしれません)。
ちなみに、沖縄本島では1967年7月に3度、米軍基地内でマダラサソリが採集されているようですが、その周辺の山地では発見されておらず (下謝名, 1972)、その後も沖縄本島でのマダラサソリの定着は知られていません。
これは、やはり南琉球と中琉球で環境の違いがあるのかもしれませんし、Kovařík et al. (2016) が述べるように、非意図的な導入個体の定着が難しいということかもしれません (もちろん、導入された個体数や雌雄にもよるでしょう)。
とにかく、日本国内でのマダラサソリの分布がケラマギャップを境にした南琉球型になっている点は非常に気になるところです。
b.海上を移動
たとえ各島が陸続きでなく海で隔てられていたとしても、サソリは海上を移動することもできると考えられます。
インドネシアのクラカタウ諸島は1883年の火山噴火により、生物がほぼ絶滅した状態となり、サソリ類についても絶滅したと考えられています。しかし、その後のサソリの分布調査では、1908年には1種、1933年には2種、1982年には3種と、記録された種数が増えており、島の外からサソリが侵入 (進入) したと考えられています (Vachon and Abe, 1988)。
Vachon and Abe (1988) は、「It seems reasonable to assume that all scorpions, as well as other animals present in the Krakatau Islands, were exterminated in 1883 when Krakatau erupted violently. Therefore, the scorpions found must have reached the Krakatau Islands by rafting from adjacent areas such as Sumatra and Java. (筆者訳:クラカタウ諸島に見られる他の動物と同様に、全てのサソリは1883年の噴火時に全滅したと考えるのが妥当だろう。なので、(調査で) 発見されたサソリはスマトラ島やジャワ島などの近隣の島から海を渡ってクラカタウ諸島へ入ってきたに違いない。)」と述べており、発見されたサソリは、それらの分布域である、クラカタウ諸島から60km離れたジャワ島や40km離れたスマトラ島から海を渡ってきたと考えています (クラカタウ諸島は、噴火時・噴火後も無人島だそうです)。
第1章で述べたカミキリムシ幼虫の海流分散が知られているように (槇原, 2013)、サソリが海を渡るとしたら、流木などに乗って流される可能性が高いでしょう。
しかし、クラカタウ諸島と比べ、先島諸島の場合、台湾と与那国島の間でも100km以上あり、海流の影響も大きく受けると思われるため、これも可能性があるとしか言えない状況です。
マダラサソリの分布:
おそらく先島諸島全域に分布している (南琉球型に一致)。
v.マダラサソリについての認識
「i.マダラサソリについての記録」で述べましたが、マダラサソリは、遅くとも1900年頃からは日本国内での生息が知られています。
今回のマダラサソリ事件での世間の反応からもわかるように、一般にはマダラサソリを在来種と考えている人が多くいると思われますが、文献等の記述から、マダラサソリについてこれまでどのように考えられてきたのかを、特定外来生物に指定された2005年の前後の時期に分けて少し整理したいと思います。
※引用箇所は全て原文のままです。
a.特定外来生物指定以前の文献
古川・高島 (1942)『南の動物』(当該箇所の執筆は高島)
「サソリは船の荷物にひそんだりして知らぬうちに甲地から乙地にと運ばれ、そこに居ついて段々ふえるといふことになりがちです。日本でも小笠原の父島や母島にもとはサソリは決して居なかつたのですが、今から十年ほど前からマダラサソリを見るやうになりました。それは日本領南洋のサイパン島から小笠原にとゞいたバナナの荷籠の中にかくれて居たのが段々ふえたのだといはれて居ます。(中略) マダラサソリは多分印度からマレー群島あたりが生れ故郷だつたのでせうが、今ではアジア・アフリカ・アメリカ等の熱帶や亞熱帶地方にひろく行きわたつて居ます。日本では琉球・臺灣・南洋諸島に産し今では小笠原に居つきになつたことは前にのベました。」
内田ら (1966)『動物系統分類学 7 (中A)』(当該箇所の執筆は江原)
「マダラサソリ Isometrus europaeus は日本にも分布し、宮古列島、八重山列島に産する。広く世界の熱帯、亜熱帯に分布する種類で、日本近くでは小笠原群島の父島、母島 (これは人為的分布であるという、牧野 1937)、台湾、内南洋などに産する」
青木 (編著) (2015)『日本産土壌動物 第二版』(当該箇所の執筆は下謝名) (1999年の初版と記述が同じなので指定前としています)
「西表島、石垣島と宮古島に分布し、主に人家周辺の日向の石下や倒木下にすむ。世界の熱帯・亜熱帯地方に広く分布。」
加納・篠永 (2003)『日本の有害節足動物』
「サソリで日本に分布しているのは、マダラサソリ Isometrus europaeus とヤエヤマサソリ Hormurus australasiae の2種で、前種は沖縄の八重山群島と小笠原諸島にかなり多数生息している。 (中略) 一方で戦前から諸外国のサソリが、船荷や木材に潜入して、日本に入り、珍しがられたり、怖がられて、ニュースとして、新聞やラジオで報道されていた。(中略) 今後国際交流がますます盛んになって、人の往来、物資の流通が増すにつれて、外国産のサソリの移入はさらに増えると思われる。このように非常にたくさんのサソリが外国から日本へ入ってきているが今日まで、これらが日本で大繁殖して帰化動物となった例はなく、セアカゴケグモとは非常に異なっている。」
今回調べたいずれの書籍でも、在来種とも外来種とも明確な記述は無いようです。「産する」「分布」「生息」という書きかたで、分布の経緯については言及がありません。
古川・高島 (1942) と内田ら (1966) は、小笠原諸島については人為分布であることを述べていますが、先島諸島についてはどちらとも書いていません。
加納・篠永 (2003) は、在来種か外来種かは記述していませんが、全体として外来サソリ問題について述べている文章中で、外国から移入されたサソリが帰化した例がないとしていることを考えると、マダラサソリとヤエヤマサソリが在来種であると考えていた可能性はあります。
外来種の問題が日本で知られるようになったのはエルトン著『侵略の生態学』(1971、原著は1958年) からとされており (自然環境研究センター, 2008)、それ以前の文献では、外来種や外来個体群であることがわかっている場合以外には、「在来である」とか「わかっていない」などをわざわざ言葉にして記述することはなかったのかもしれません。少なくとも、外来種問題が知られるようになった以降では、外来サソリについて記述している一方で、マダラサソリが外来種である可能性には全く触れていないことから、外来種とは考えられていなかったのではないかと推測されます。
b.特定外来生物指定以降の文献等
(マダラサソリ事件後に発行されたものは事件の影響を受けていると思われるため省略)
池田 (監修) (2006)『外来生物事典』
「サソリが荷物に混入して、国内の港で発見され大騒ぎになることがあるが、外来のキョクトウサソリ科が野生化した事例は報告されておらず、定着も確認されていない。国内には中型のキョクトウサソリ科マダラサソリが八重山諸島、宮古諸島、小笠原諸島に定着している。」
相原・秋山 (2007)『節足動物ビジュアルガイド タランチュラ&サソリ』
「日本を含め人の移動に付随して棲息域を広めたとされる種で、開けたところにある倒木の下、岩場、積み上げられた板の隙間などから見つかり、森林の中ではあまり見つからない。Buthidaeの中で、本種だけは江戸時代以前から沖縄に帰化していたため特定外来生物の指定を受けなかったので、現在でも飼養は可能。」
「例外は、東南アジアに元々棲息していて、流木や船に乗って数百年前に南西諸島に帰化したといわれるマダラサソリぐらいだ。」
自然環境研究センター (編著) (2008)『日本の外来生物』(当該箇所の執筆は石塚・岸本)
「宮古・八重山諸島に生息し、小笠原からの記録もあるマダラサソリ Isometrus maculatus は本科に属するが、宮古・八重山諸島においては在来種もしくは明治時代より前の外来種と考えられるため、特定外来生物から除外される。」
石川 (編) (2008)『節足動物の多様性と系統』(当該箇所の執筆は小野)
「日本にはマダラサソリ Isometrus europaeus (キョクトウサソリ科。宮古、八重山諸島、小笠原) と、ヤエヤマサソリ Liocheles australasiae (ヤセサソリ科。宮古、八重山諸島) の2種を産する。貨物などについて人為的に移入された外来種がときどき発見されるが、定着した例はない。」
「人体に有毒な種があり、日本ではキョクトウサソリ科全種が特定外来生物に指定されている。」
小野 (編著) (2009)『日本産クモ類』(当該箇所の執筆は小野)
「日本にはマダラサソリ Isometrus europaeus (キョクトウサソリ科) (宮古,八重山諸島,小笠原) と、ヤエヤマサソリ Liocheles australasiae (ヤセサソリ科) (宮古,八重山諸島) の2種を産する。貨物などについて人為的に移入された外来種がときどき発見されるが、定着した例はない。人体に有毒な種があり、在来種を除くキョクトウサソリ科全種が特定外来生物に指定されている。」
国立環境研究所 「侵入生物データベース:キョクトウサソリ科」 (2012更新)
自然分布:約89属950種が,アフリカ,中米,アジア,オーストラリアなど熱帯・亜熱帯・温帯の一部に広域分布。日本国内では,先島諸島 (宮古島,石垣島,西表島) に広域分布種マダラサソリ (Isometrus europaeus) が分布。
国内移入分布:外来のものは国内に定着していない。広域分布種マダラサソリ Isometrus europaeus が八重山諸島 (宮古島,石垣島,西表島) にも自然分布。
池田 (2006) は、外来生物法を前提とした書籍であるためだと思われますが、「定着」という言葉を使用しており、国外から侵入 (進入) したという認識となります。マダラサソリが国内に入ってきた理由について明言はしていませんが、外来のサソリが定着したことはないとしているため、マダラサソリが導入による外来種とは考えていないと言ってよいかもしれません。
相原・秋山 (2007) は、「日本を含め人の移動に付随して棲息域を広めた」「流木や船に乗って帰化した」としており、マダラサソリは外来種という認識であることがわかります (もし流木に乗って国内に入ってきたのであれば、分散による在来種と言えるでしょうが)。
自然環境研究センター (2008) は「在来種もしくは明治時代より前の外来種と考えられる」としており、どちらの可能性もある、つまりどちらかわからないという認識になります。また、2019年の改訂版では、キョクトウサソリ科の項目の全体的な記述内容は同じですが、マダラサソリについての記述だけが削除されています。マダラサソリ事件後となっては、「特定外来生物から除外される」という記述は明らかな間違いであるため削除されるべきですが、日本国内にマダラサソリが分布していること自体が削除されると、国内にはキョクトウサソリ科のサソリが分布していないと読者に誤解を与えるおそれがあります。
石川 (2008:執筆は小野) や小野 (2009) は、外来種が定着した例はないとしています。また、小野 (2009) は、はっきりとは書いていませんが、キョクトウサソリ科のマダラサソリが国内に生息していると述べたうえで、「在来種を除くキョクトウサソリ科全種が特定外来生物に指定されている」と述べていることから、国内に生息するキョクトウサソリ科であるマダラサソリは在来種であるという認識だと解釈できます。
国立環境研究所「侵入生物データベース」では、キョクトウサソリ科のサソリは「今後日本への導入・定着が起きないよう特に注意すべき」リストに挙げられおり、「外来のものは国内に定着していない」「自然分布」という記述から、マダラサソリが在来種であるとの認識であると考えられます。
いずれの文献でも、はっきりとマダラサソリについて「外来種」「在来種」という言葉で記述しているものはありませんでしたが、多くの著者は、外来サソリ類やキョクトウサソリ科についての記述から、マダラサソリを在来種と認識していたと解釈するのが妥当でしょう。上記 (ii) の全てでキョクトウサソリ科全種が特定外来生物に指定されていることを記述しているので、日本国内に生息するキョクトウサソリ科であるマダラサソリについて在来種か外来種か (の認識) を明確にすべきであったと思うのですが、なんとなく在来種と思えるようなか書きかたをするだけで、あいまいにしているという印象です (ただし、間違ってはいますが、相原・秋山, 2007 や自然環境研究センター, 2008 が、マダラサソリが特定外来生物になるかどうかをはっきり述べていることは大切です)。
なぜこのようなあいまいな書きかたがされているのかを考えてみると、相原・秋山 (2007) や自然環境研究センター (2008) に見られるような、マダラサソリが古くから国内に分布しているサソリだという認識が大きく影響していたのではないでしょうか。
マダラサソリという和名が命名された岩川 (1906) の論文タイトルが『本邦産の蝎類』、つまり「日本のサソリ」という意味ですし、マダラサソリの分布している台湾が当時既に日本統治下にあり、さらに第一次世界大戦後、いわゆる南洋諸島が日本の委任統治領となって、多くの島々でマダラサソリが見つかったことで (江崎, 1943)、(サソリについて知る) 日本人のなかでは「マダラサソリは日本のサソリ」という認識が広まっていったと考えられます。上述のとおり、外来種問題が日本でも知られるようになったのは『侵略の生態学』(エルトン, 1971) からであり (自然環境研究センター, 2008)、それ以前から日本のサソリと考えられてきたマダラサソリについては、なんとなく在来種だろうと考えられてきたというのが実際のところではないでしょうか (なので、キョクトウサソリ科全種が特定外来生物という状況になって、今まで在来種と考えていたマダラサソリについてどう書いていいかわからないとなったのかもしれません)。
マダラサソリについての認識:
古くから国内に分布しており、なんとなく在来種だろうと認識されてきた (と考えるのが妥当)。
マダラサソリは外来種と考えられるか?
ここまで見てきた外来種判定のポイントをまとめると、以下のようになります。
マダラサソリは、先島諸島全域に分布していると考えられる。最も古い記録は1906年で、外来種問題が知られる前から日本国内に分布しているため、なんとなく在来種だろうと認識されてきた。
総合的には「在来種>外来種」とも言えそうですが、輸入された記録や化石記録、遺伝的解析の研究などの直接的なデータが無いため、はっきりとは言い切れないというのが結論です。
では、なぜはっきりと断言できる証拠のようなものが無いかというと、結局のところ、これまでマダラサソリが問題にならなかったからだと思います。在来種か外来種かはっきりわかっていない生物のなかで、ハクビシンのように被害が出て問題になるものは研究が行われますが、そうでない場合、外来種の可能性を疑い調べる必要はなかったでしょう。だから、いきなりマダラサソリは特定外来生物ですと言われても、在来種である証拠がない (と同時に外来種である証拠もない) となるのです。
今後もしかすると、マダラサソリの研究が進められどちらか結論が出るかもしれません。
しかし、上述の通り、在来種であろうと外来種であろうと、20世紀初頭に「マダラサソリ」という和名が命名されたときからマダラサソリは「日本のサソリ」と考えられ、親しまれてきた種であることに違いはありません。そして、マダラサソリはこれまで問題になったことがなく、特定外来生物の定義にあるような、被害を及ぼすおそれのある動物ではないと言えます。
出典 ▼
- 岩川友太郎 (1906) 本邦産の蝎類, 動物学雑誌, 18(207):5-12.
- 黒岩恒 (1899) 石垣島 (承前), 地質学雑誌, 6(72):307-311.
- 黒岩恒 (1899) 石垣島, 地質学雑誌, 6(71):283-289.
- 高島春雄 (1943) 蠍襍記, Acta Arachnologica, 8(3):58-66.
- H. Takashima (1954) Considerations on the change of animal life in Japan (II), Journal of the Yamashina Institute for Ornithology, 1(4):146-155.
- 古川晴男, 高島春雄 (1942) 南の動物, 光風館, 東京.
- 環境省 (2007) 小笠原の自然環境の保全と再生に関する基本計画.
- 新たな外来種の侵入・拡散防止に関するワーキンググループ (2013) 世界遺産地域 小笠原諸島 新たな外来種の侵入・拡散防止行動計画の策定に向けた課題整理.
- 久保快哉 (2001) 硫黄島訪問記, やどりが, 2001(188):64-68.
- 相原和久, 秋山智隆 (2007) タランチュラ&サソリ (節足動物ビジュアルガイド), 誠文堂新光社, 東京.
- 青木淳一 (編著) (2015) 日本産土壌動物: 分類のための図解検索 第2版, 東海大学出版部, 秦野.
- 下謝名松栄 (1972) 琉球列島の蛛形類の分布, 遺伝, 26:100-106.
- 山﨑仁也, 松村雅史, 吉田和久, 力身恭二, 目黒賢児 (2016) 鳩間島・新城(上地)島・黒島の動物相(FAUNA)~昆虫相を中心に~, 鳩間島・新城島・黒島総合調査報告書, 沖縄県立博物館・ 美術館博物館, 那覇.
- Fet, V., WD. Sissom, G. Lowe, and ME. Braunwalder (2000) Catalog of the scorpions of the world (1758-1998), The New York Entomological Society, New York.
- Di, ZY., ZZ. Yang, SJ. Yin, ZJ. Cao and WX. Li (2014) History of study, updated checklist, distribution and key of scorpions (Arachnida: Scorpiones) from China, Zoological research, 35(1):3–19.
- Di, ZY., ZJ. Cao, YL. Wu, L. Zhu, H. Liu and WX. Li (2013) The scorpions of Hainan Island, China (Arachnida: Scorpiones), Euscorpius, 153:1-22.
- Sulakhe, S., N. Dandekar, A. Padhye, and D. Bastawade (2020) Two new cryptic species of Isometrus (Scorpiones: Buthidae) from the northern Western Ghats, India. Euscorpius, 305:1-24.
- Kovařík, F. (2003) A review of the genus Isometrus Ehrenberg, 1828 (Scorpiones: Buthidae) with descriptions of four new species from Asia and Australia, Euscorpius, 10:1-19.
- Kovařík, F., G. Lowe, KB. Ranawana, D. Hoferek, VS. Jayarathne, J. Plíšková and F. Šťáhlavský (2016) Scorpions of Sri Lanka (Scorpiones: Buthidae, Chaerilidae, Scorpionidae) with description of four new species of the genera Charmus Karsch, 1879 and Reddyanus Vachon, 1972, stat. n., Euscorpius, 220:1-133.
- 琉球大学 (2019) 南琉球最古の土器の謎を解明 ~新たな非破壊的な理化学分析で明らかになった先史土器文化の変遷~, 琉球大学ウェブサイト. (https://www.u-ryukyu.ac.jp/news/6760/), (2020/05/07参照)
- Lourenço, WR and D. Huber (2002) New addition to the scorpion fauna (Arachnida: Scorpiones) of Sri Lanka, Revue suisse de zoologie, 109(2):265-275.
- Pocock, RI. (1900) Arachnida, Fauna of British India, including Ceylon and Burma, Taylor and Francis, London.
- Wallace, AR. (1876) The geographical distribution of animals, with a study of the relations of living and extinct faunas as elucidating the past changes of the earth's surface, Harper & Brothers, New York.
- 木村政昭 (1996) 琉球弧の第四紀古地理, 地学雑誌, 105(3):259-285.
- 諸喜田茂充 (1996) 琉球列島の河川陸封コエビ類とサワガニ類の起源論, 地学雑誌, 105(3):343-353.
- 池安間繁樹 (2001) 琉球列島: 生物の多様性と列島のおいたち, 東海大学出版会, 東京.
- 河村善也 (2014) 日本とその周辺の東アジアにおける第四紀哺乳動物相の研究—これまでの研究を振り返って—, 第四紀研究, 53(3):119-142.
- 宮城邦治, 小倉剛, 伊澤雅子 (2002) 「琉球列島の動物たち—特異な動物相の成り立ちを探る—」企画趣旨と報告 (大会シンポジウム報告), 哺乳類科学, 42(1):85-91.
- 環境省 (2010) 奄美・琉球諸島の生物多様性 島々に棲む様々な生きものたち.
- Vachon, M. and T. Abe (1988) Colonization of the Krakatau Islands (Indonesia) by scorpions, Acta Arachnologica, 37(1):23-32.
- 槇原寛 (2013) 移動する森林昆虫類 (1) 小笠原諸島のカミキリムシ類, 海外の森林と林業, 88:58-62.
- 内田亨 (監修) (1966) 動物系統分類学 7 (中A), 中山書店, 東京.
- 加納六郎, 篠永哲 (2003) 新版 日本の有害節足動物 生態と環境変化に伴う変遷, 東海大学出版会, 秦野.
- チャールズ・S・エルトン, 川那部浩哉 (訳), 大沢秀行 (訳), 安部琢哉 (訳) (1971) 侵略の生態学, 思索社, 東京.
- 多紀保彦 (監修), 自然環境研究センター (編著) (2008) 決定版 日本の外来生物, 平凡社, 東京.
- 池田清彦 (監修), DECO (編) (2006) 外来生物事典, 東京書籍, 東京.
- 岩槻邦男 (監修), 馬渡峻輔 (監修), 石川良輔 (編) (2008) 節足動物の多様性と系統 (バイオディバーシティ・シリーズ 6), 裳華房, 東京.
- 小野展嗣 (編著) (2009) 日本産クモ類, 東海大学出版会, 秦野.
- 国立環境研究所. "キョクトウサソリ科". 侵入生物データベース. (https://www.nies.go.jp/biodiversity/invasive/DB/detail/70070.html), (2020/05/07参照)
- 自然環境研究センター (編著) (2019) 最新 日本の外来生物, 平凡社, 東京.
- 江崎悌三 (1943) 南洋群島の蠍, Acta Arachnologica, 8(1-2):1-5.
